
仙人掌科(Cactaceae)是被子植物门木兰纲石竹目的一个科,一般统称为仙人掌,大约包含有139属,大约1500至1800种植物。该科的大多数种类分布于美洲,以墨西哥及中美洲为分布中心;但也有例外,丝苇属(Rhipsalis)不仅分布于美洲,在热带非洲、马达加斯加和斯里兰卡等地也均有发现。
形态特征
编辑仙人掌科植物为多年生肉质草本、灌木或乔木,地生或附生。根系浅,开展,有时具块根。攀爬、匍匐和附生类的仙人掌可能只有不定根,常见于茎与生根介质接触的地方。
原产于墨西哥的金琥(Echinocactus grusonii),世界各地广泛栽培


肉质茎
大多数仙人掌的茎是绿色的,通常为蓝绿色或棕绿色。茎中含有叶绿素和气孔,能够进行光合作用。此外仙人掌茎通常呈明显的蜡质。茎常肉质,适合储存水分;直立、匍匐、悬垂或攀援,圆柱状、球状、侧扁或叶状;节常缢缩,节间具棱、角、瘤突或平坦,具水汁,稀具乳汁;小窠螺旋状散生,或沿棱、角或瘤突着生,常有腋芽或短枝变态形成的刺,稀无刺,分枝和花均从小窠发出。茎表面光滑(如仙人掌属的某些物种)或覆盖着各种突起。乳突球属(Mammillaria)植物的突起变化较多,从小的隆起到较大的乳头状凸出;岩牡丹属(Ariocarpus)植物的突起形似叶片。茎呈肋状或凹槽状,其突出程度取决于茎内储存的水分含量。当茎中充满水时(仙人掌的含水量可高达质量的90%),茎肿胀而看不出呈肋状;而当缺水时茎萎缩,明显呈肋状。

园艺栽培的丝苇属某种
小窠
茎上萌发刺或花的多毛区域称为小窠,是仙人掌植物的特有结构,其形态变化较大。小窠是高度特化和浓缩的芽或枝。在正常的枝条中,带有叶子或花朵的节点间存在一定距离,而在小窠中,节点非常接近,从而形成独立的结构。小窠为圆形或椭圆形,或分为相互连接的两部分(如通过茎中的凹槽),或完全分开(二形小窠)。靠近茎顶部的小窠常产生花朵,其他部分则产生刺。小窠通常具有毛状体,因此外观常呈毛状,有时具有独特的颜色,如黄色或棕色。

在大多数仙人掌中,小窠仅在几年内产生新的刺或花朵,然后其活性降低,刺的数量相对固定。在与仙人掌祖先类群亲缘关系较近的木麒麟属(Pereskia)、仙人掌属(Opuntia)和大冠柱属(Neoraimondia)中,小窠保持活性的时间要长得多。
叶片
叶扁平,全缘或圆柱状、针状、钻形至圆锥状,互生,或完全退化,无托叶。绝大多数仙人掌没有可见的叶子,其光合作用发生在茎中。除了木麒麟属(Pereskia)和蔷薇麒麟属(Rhodocactus)等,它们形如普通树木或灌木,有许多叶子,这些叶子具有中脉且两侧扁平。仙人掌组(仙人掌亚科)中的种类也有可见的叶子(如麒麟掌属Pereskiopsis),宿存或仅在生长季节产生然后脱落。卧麒麟属(Maihuenia)可依靠叶子进行光合作用。有的仙人掌叶片非常小,肉眼近不可见,很多种类叶片小于1.5毫米,有约一半种类小于0.5毫米。但是,这种叶片不能进行光合作用,有学者提出这种叶片可能在植物激素的产生和腋芽确定方面发挥一定的作用。

长有许多叶片的木麒麟
特化的刺
仙人掌上的刺是特化的叶片,来源于小窠。有的有叶仙人掌上也存在刺,例如木麒麟属、麒麟掌属和卧麒麟属,这说明仙人掌在叶片完全进化前,就已经出现了刺。有些仙人掌只在幼苗时有刺,尤其是一些附生性的仙人掌,如丝苇属(Rhipsalis)和仙人指属(Schlumbergera),或一些地生仙人掌(如岩牡丹属)。仙人掌的刺是物种鉴定的重要依据,其数量,颜色,大小,形状和硬度以及小窠上的刺等特征,存在较为明显的种间差异。大多数仙人掌具直刺,有的种类的刺则略微弯曲。一些仙人掌刺比较扁平(如月童Sclerocactus papyracanthus)。另外,有的仙人掌有钩刺(如海泉Mammillaria rekoi)具有一个或多个钩状的中央棘和直的外刺。除了正常长度的刺外,仙人掌亚科的成员还具有相对较短的刺,容易脱落,称为钩刺。这些刺进入皮肤后,由于其非常细且容易破裂,而难以去除,造成长时间刺激。
花
花白、黄、红或洋红,常单生,少数如木麒麟属的花簇生。无梗,稀具梗并组成总状、聚伞状或圆锥状花序,两性花,稀单性花,辐射对称或左右对称;花托通常与子房合生,稀分生,上部常延伸成花托筒,外面覆以鳞片(苞片)和小窠,稀裸露(如裸萼球属Gymnocalycium),小窠内可产生毛或刺;花被片多数和无定数,螺旋状贴生于花托或花托筒上部,外轮萼片状,内轮花瓣状,或无明显分化;雄蕊多数,着生于花托或花托筒内面中部至口部,螺旋状或排成两列;花药基部着生,2室,药室平行,纵裂。雄蕊基部至子房之间常有蜜腺或蜜腺腔。雌蕊由3至多数心皮合生而成;子房通常下位,稀半下位或上位,1室,具3至多数侧膜胎座,或侧膜胎座简化为基底胎座状或悬垂胎座状;胚珠多数至少数,弯生至倒生;花柱1,顶生;柱头3至多数,不分裂或分裂,内面具多数乳突。

开花的裸萼球属植物绯花玉(Gymnocalycium baldianum)

在夜晚开花的昙花(Epiphyllum oxypetalum)
果实和种子
浆果肉质,常具黏液,稀干燥或开裂,散生鳞片和小窠,稀裸露。种子多数,稀少数至单生;种皮坚硬,有时具骨质假种皮和种阜,无毛或被绵毛;胚通常弯曲,稀直伸;胚乳存在或缺失;子叶叶状扁平至圆锥状。

售卖时被称为红心火龙果的哥斯达黎加量天尺(Hylocereus costaricensis)

中国广西的火龙果种植田,用做水果食用的仙人掌科植物量天尺果实,通常和其他几种仙人掌科植物果实被统称为火龙果
适应性
编辑大多数仙人掌科植物,非常适合生存于炎热和干燥的环境(旱生植物),可以通过采取一些特别的措施,促进自身更高效的水资源利用。

中国厦门栽培的仙人掌
特化的叶和刺
叶片保水的关键在于表面积与体积的比率。失水速率与表面积成正比,而水的保有量与体积成正比。与表面积体积比低的结构(如仙人掌粗大的茎)相比,表面积体积比高的结构(一般植物薄薄的叶片)会以更高的速率失去水分。为适应炎热、干燥的环境,大多数仙人掌没有明显的叶片,只有少数种类,如木麒麟属的部分物种,与仙人掌的祖先类群较为接近,都有肥厚且多汁的叶片;麒麟掌属也有肉质的叶片。
仙人掌的刺由叶片特化而来,长叶片的仙人掌也有刺,这说明在仙人掌完全失去叶片之间,就已经进化出了刺。虽然刺具有较高的表面积/体积比,但其由死细胞纤维构成,在成熟时,含水量很少甚至不含水。刺可以让空气聚集在仙人掌表面,形成湿润的保护层,从而降低蒸腾作用。并为植株遮挡阳光,降低仙人掌表面的温度、减少了水分的流失。当空气足够潮湿时,比如在大雾或清晨的薄雾中,刺上会凝结水分,然后滴落到地面上,被根部吸收。这些刺不仅在帮助植物保持水分,还为一些物种,提供了保护和伪装。

仙人掌坚硬的刺
肉质茎
大多数仙人掌的茎是肉质的,是其主要贮存水分的器官。水分大约占到仙人掌总质量的90%。不同种类的仙人掌,茎的形状差别巨大。柱状仙人掌、球形仙人掌的茎具有较低的表面积/体积比,可以减少水分的损失,也将阳光的加热效应降至最低。许多仙人掌的茎带棱或凹槽,在干旱时期收缩,在水分充足时吸水膨胀。一棵成熟的仙人掌,据说可以在一场暴雨中吸收多达760升的水。茎的外层通常有一个坚韧的蜡质层,可以减少水分流失,因此许多仙人掌的茎部颜色偏灰或偏蓝,大多数仙人掌的茎具有叶绿体,可以在没有叶子的情况下进行光合作用。
广且浅的根系
许多仙人掌的根伸展得很广,但分布较浅。有报道称,一株年幼的仙人掌只有12厘米高,但根系分布范围直径可达2米,但深度不超过10厘米。旱地降雨时,仙人掌能迅速长出新根。仙人掌根细胞液的盐浓度也相对较高,这些都使得仙人掌能够在降水时迅速吸收水分。龙爪球(Copiapoa atacamensis)生长于世界上最干燥的地方之一的智利北部的阿塔卡马沙漠,主根长度可能是地表植株的几倍长。

生长于智利干旱沙漠中的龙爪球属植物
景天酸代谢(CAM)
光合作用需要植物吸收二氧化碳,同时通过蒸腾作用失去水分。仙人掌可以通过进行光合作用来减少水分的流失。大部分植物利用C3机制,即白天,树叶内部的空气中的二氧化碳不断被吸收,首先转化3-磷酸甘油酸酯,然后转化为碳水化合物等产物。气孔可以打开和关闭,从而控制空气进入植物内部空间。在光合作用期间需要持续的二氧化碳供应,这意味着气孔必须是开放的,因此水蒸气会持续流失。使用C3机制的植物以这种方式失去了97%从根部吸收的水分。更进一步来说,随着温度的升高,捕获二氧化碳的酶反而开始捕获越来越多的氧气,从而使光合作用的效率降低了多达25%。
景天酸代谢(CAM)是仙人掌等多肉植物,为避免C3机制带来的失水问题,而进化出的代谢机制。在景天酸代谢过程中,气孔只在夜间开放,此时温度和水分损失最低。二氧化碳进入植物,以有机酸的形式被捕获并储存在细胞内(液泡中)。气孔白天时处于关闭状态,光合作用只利用这些晚上储存的二氧化碳。景天酸代谢能够更有效地利用水,但代价是减少了从大气中固定的碳量,从而可用于生长的较少。
为了进行光合作用,仙人掌的茎经历了多次改变。在进化早期,大部分现生的仙人掌(海麒麟属除外)的祖先类群,茎上发育出气孔,树皮发育延迟。并且,茎发育出通常只在叶片中存在的类似结构,外表皮之下产生细胞壁增厚的细胞,提供机械支撑。细胞之间形成了通气组织,让二氧化碳可以向内部扩散。茎中心的皮层,发育出一种由含有叶绿体的相对非特化细胞组成的植物组织(绿色组织),排列成海绵状层和栅栏状层(大部分光合作用都发生于此)。
分布范围
编辑仙人掌科主要产于美洲,分布范围从南美洲安第斯山脉以东的巴塔哥尼亚延伸到加拿大西部。物种存在许多个多样性中心,适应干旱环境的仙人掌,主要存在三个多样性中心:墨西哥和美国西南部、安第斯山脉西南部(秘鲁、玻利维亚、智利和阿根廷)以及巴西东部(远离亚马孙盆地的区域)。而附生在树木或攀援的仙人掌,需要潮湿的环境,因此与旱生仙人掌具有不同的多样性中心,主要分布在巴西东南部沿海山区和大西洋沿岸森林中。
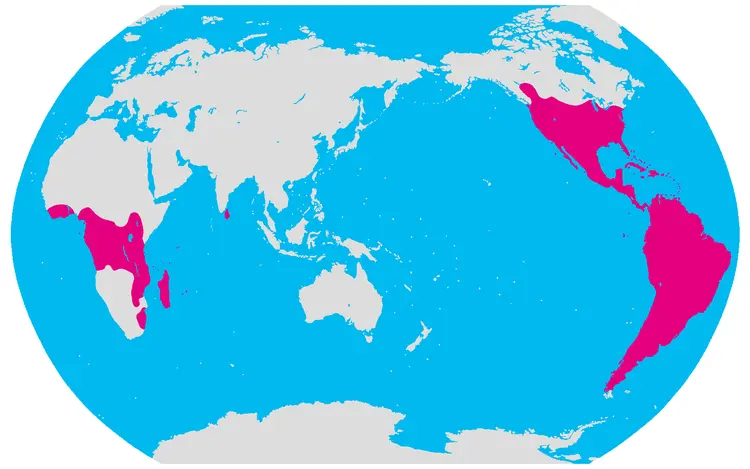
仙人掌科的分布图
丝苇属是个例外,其不仅原产于美洲,还在热带非洲、马达加斯加和斯里兰卡都有发现。有一种理论认为,它是通过种子被候鸟取食后传播的,美洲以外的种群是多倍体,被视为不同的亚种,这种现象支持了该物种传播历史较早的观点。而另一种理论认为,该物种最初是通过南美洲和非洲之间贸易的船只,穿越大西洋,然后由鸟类取食进而产生了更广泛的传播。
传播入侵
编辑关于人类第一次踏入有仙人掌分布的美洲区域的确切、以及首次使用仙人掌的时间,一直存在争议。根据智利的一处考古遗址,人类发现仙人掌的历史,最早可以追溯到大约1.5万年前。在巴西卡皮瓦拉山脉(Serra da Capivara)的洞穴壁画中,发现了仙人掌的图案;墨西哥、秘鲁的古代垃圾堆中,发现了仙人掌种子,时间大约在1.2万至9000年前,这可能是当时的采集者,在野外采集仙人掌果实,带回营地后所遗留下的。

用于制作胭脂红的胭脂虫感染的仙人掌
在15世纪末,欧洲人到达美洲时第一次见到了仙人掌。第一次登陆的西印度群岛时,欧洲人发现的仙人掌属植物较少,如花座球属(Melocactus)。因此,花座球属可能是欧洲人最早发现的仙人掌植物。花座球属也在16世纪末前,出现在英国的仙人掌收藏中,被称为Echinomelocactus,到了18世纪早期,该名称由植物学家将学名缩短为Melocactus,也就是花座球的属名。随后,一些仙人掌的观赏和可食用的种类陆续被引入到欧洲。因此在1753年时,瑞典博物学家、生物命名之父卡尔·林奈(Carl Linnaeus)得以命名了22种仙人掌科植物,在一个名为Cactus opuntia的植物中,有这样的描述:“大果……在西班牙和葡萄牙”,表明其在欧洲早期有被使用的历史。
在中国,仙人掌在明朝末年作为围篱被引种,南方地区也常见有栽培。梨果仙人掌在原产地墨西哥的栽培历史很长,在史前时期就已经被人类驯化,野生种和栽培种的分布边界模糊,后被作为观赏植物、饲料、果树大范围引至世界各地。16世纪,梨果仙人掌首次从美洲引入到了欧洲,1645年又由荷兰人首次引入中国台湾栽培,中国南方各地作为围篱引种。1702年的《岭南杂记》中,首次记载了仙人掌。2002年,根据中国学者李振宇和解焱报道,仙人掌在中国广东、 香港、澳门、广西南部、海南沿海地区及南海诸岛均逸为野生,随后仙人掌被多次报道在多个省份入侵。17 世纪中期,单刺仙人掌(Opuntia monacantha)作为果树和饲料作物被引至世界各地。根据明朝刘文征1625年所著的《滇志》,在当时,单刺仙人掌已经在云南被当做花卉引种和栽培。
清代吴其濬所著的《植物名实图考》中所描绘的仙人掌
分类演化
编辑早期分类
仙人掌科的学名为Cactaceae,由该科的模式属乳突球属学名Mammillaria的异名Cactus的复合形式Cact-,加上表示科的等级后缀-aceae构成。
自第一株仙人掌被人类发现并描述以来,其命名和分类就存在诸多困难和争议。1737年,卡尔·林奈(Carl Linnaeus)将其所知的仙人掌分成了两个属,原仙人掌属(Cactus)和木麒麟属(Pereskia)。然而,在1753年出版《植物种志》时,他又把它们都归入到同一个属——原仙人掌属(Cactus)中。“Cactus”一词源自拉丁语κάκτος(kaktos),是古希腊人泰奥弗拉斯图斯用于多刺的植物上的名称。
后来的植物学家,如1754年的苏格兰植物学家菲利普·米勒(Philip Miller)则将仙人掌分为可几个不同的属;1789年,法国植物学家安托万·洛朗·德·尤西厄(Antoine Laurent de Jussieu)又将这些属放入他新创建的仙人掌科(Cactaceae)中。
科名确立
20世纪初,植物学家们认为林奈所命名的“Cactus”一词意义混乱,无法确定究竟是属名还是科名。因此,在1905年举行的维也纳植物学大会上,集体决议不再采用“Cactus”这个名称,宣布以Mammillaria(乳突球属)作为仙人掌科的模式属,保留Cactaceae(仙人掌科)的名称,也导致了仙人掌科中,不包含以其科名(Cactaceae)来命名的属(Cactus)这一特殊情况。

仙人掌科的模式属乳突球属某种
而该科的中文名“仙人掌科”与模式属“乳突球属”的中文名不一致,是因为该科的中文名源于日本学者拟定的日文科名,当时的学者选用了更知名的栽培属仙人掌属(Opuntia)为该科命名,随后这一日文科名又被译为了中文科名。
命名困难
植物的命名依赖于模式标本,这也是仙人掌科分类命名的难点之一。模式标本通常需要压缩和干燥,然后储存在标本馆中以作为参考凭证。但是仙人掌的形态特殊,标本制作困难。除此以外,许多仙人掌是栽培者和园艺家来命名的(而非专业的植物学家),命名时并没有遵循植物命名的专业规范(即《藻类、真菌和植物的国际命名法规》中的相关规定)。例如,德国园艺学家柯特·巴克伯格(Curt Backeberg)就命名或重命名了多达1200个类群,且从未附上模式标本,因此有学者表示这种情况“留下了一连串的命名混乱,可能会困扰仙人掌分类学家几个世纪”。
亚科划分
1984年,国际多肉植物研究组织为仙人掌科单独成立了学术工作小组,即国际仙人掌科系统学小组(ICSG),以深入建立仙人掌科在属一级的分类系统共识。他们的系统也为随后仙人掌科科下划分的奠定了基础。21世纪所发表的研究,将仙人掌分为约125至140个属,1500至1800个种,排列成许多族和亚科。2001年,ICSG共承认木麒麟亚科、仙人掌亚科、卧麒麟亚科、仙人柱亚科等四个亚科。2015年,一项研究将海麒麟属从木麒麟亚科中分出,并建立了一个新亚科——海麒麟亚科。
- 木麒麟亚科(Subfam. Pereskioideae)
ICSG分类中,该亚科只有唯一的广义木麒麟属,其形态接近于仙人掌科祖先,植株为有叶片的乔木状或灌木状;茎横截面光滑,呈圆形,而不是像其他仙人掌一样呈肋状或有结。其光合作用可以使用两类系统,C3机制和景天酸机制(CAM)。分子系统发育研究表明,广义的木麒麟属(Pereskia)不为单系,后被分为三个属,即海麒麟属(2015年独立成海麒麟亚科),蔷薇麒麟属(Rhodocactus)和狭义的木麒麟属(Pereskia)。

木麒麟属的大叶木麒麟(Pereskia grandifolia)
- 海麒麟亚科(Subfam. Leuenbergerioideae)
海麒麟属(Leuenbergeria)原归属于木麒麟亚科,被单独置于一个单独的单属亚科海麒麟亚科(Leuenbergerioideae)中。
- 仙人掌亚科(Subfam. Opuntioideae)
该亚科包括约15个属,植株幼时可能有叶子,成年后消失,茎通常分为不同的关节或“扁平的叶状茎。植株大小差异大,有从小垫状的卧云掌属(Maihueniopsis),到高达10米甚至更高的树状的仙人掌属(Opuntia)。
- 卧麒麟亚科(Subfam. Maihuenioideae)
该科唯一的属为卧麒麟属(Maihuenia),该属内有两种,均可为低矮垫状,具有仙人掌的一些原始特征(植物有叶片,无景天酸代谢)。
- 仙人柱亚科(Subfam. Cactoideae)
该亚科是最大的亚科,共分为九个族,包括所有一般被认为是仙人掌的典型种类。亚科内成员的习性变化大,从树状到附生均有。通常没有叶片,且茎不成节,呈肋状或突起状。其中的两个族,量天尺族(Hylocereeae)和丝苇族(Rhipsalideae),包含外观与典型仙人掌不同,为攀援或附生形式,具有扁平的茎。

仙人柱亚科
进化历史
仙人掌科没有发现可以显示其进化历史的化石,但是地理分布可以为其进化历史提供一些线索和证据。仙人掌产于南美洲和北美南部地区,表明该科是在古代冈瓦纳大陆分裂为南美洲和非洲之后进化而来的,大约发生在1.45~1.01亿年前的白垩纪早期。仙人掌在大陆分裂之后,具体何时进化的并不清楚。较早的资料表明,它的早期起源大约在9000万至6600万年前,即白垩纪晚期。一些分子研究所表明的起源时间更晚一些,可能是在始新世晚期至渐新世早期,大约在3500万至3000万年前。
根据仙人掌的系统发育研究,最早的分化类群(海麒麟属)可能起源于中美洲和南美洲北部,而具有肉质茎的仙人掌(core cacti)则是后来在南美洲南部进化出来的,然后向北迁移。具有明显肉质茎的仙人掌,估计在大约2500万年前就已经进化了。其进可能是由于大约2500万至2000万年前,安第斯山脉中部发生了隆起,与干旱的增加和变化有关。而仙人掌的物种多样性增高,是在1000至500万年前(中新世晚期到上新世)才出现的。其他多肉植物,如南非的番杏科(Aizoaceae),马达加斯加的刺戟木科(Didiereaceae)和美洲的龙舌兰属(Agave),可能也在同一时期多样化增高,与全球干旱环境面积扩大化相吻合。
代表类型
编辑乔木状仙人掌
乔木仙人掌具有一个或多个的木质主干和枝干。海麒麟属、木麒麟属和蔷薇麒麟属中的植物,枝干被叶片覆盖,常常不被当做是仙人掌。大多数乔木仙人掌中更典型的特征,是树干上几乎没有任何叶片和树皮,且覆盖着小刺,如武伦柱(Pachycereus pringlei)和其他大型仙人掌属植物。一些仙人掌大小可能和普通的树木一样,但没有树枝,如大型的广刺球(Echinocactus platyacanthus)。仙人掌也有灌木状的种类,如大王阁(Stenocereus thurberi)。

栽培的广刺球,自然条件下可以长成粗大的乔木状
柱状仙人掌
柱状仙人掌由直立的圆柱形茎组成,分支或不分支,无明确的主干和分支。柱状仙人掌与乔木或灌木之间的界限比较模糊。例如,年幼矮小的翁柱(Cephalocereus senilis)个体为柱状,而年老高大的标本可能会变成树状。

翁柱(Cephalocereus senilis),又称白头翁
球状仙人掌
球状仙人掌具有比柱状仙人掌,更短且呈球形的茎。球状仙人掌可能是独立生长的,如日之出丸(Ferocactus latispinus),也可能簇生,每簇中的所有或部分主干可能拥有共同的根。

簇生的仙人掌
其他类型
其他仙人掌的外观存在较大差异。在热带地区,有些仙人掌以攀援或附生的形式生长,它们通常具有扁平且形似叶片的茎、少刺甚至无刺。有的攀援类的仙人掌体型非常大,据报道量天尺属(Hylocereus)的一个标本从根到最远的茎长达100米。附生类的仙人掌,如丝苇属或仙人指属(Schlumbergera)的物种,常生长在高出地面的树木中,下垂形成密集的团块。

仙人指属常见的园艺植物蟹爪兰(Schlumbergera truncata)
繁殖
编辑自然状态下,大部分仙人掌的花是由昆虫、鸟类或蝙蝠进行授粉,极少数物种会发生自花授粉,如天惠球属(Frailea)的某些植物,其花不开放,进行闭花受精。由蝴蝶授粉的花通常颜色鲜艳,白天开放,而由飞蛾授粉的花通常颜色为白色或浅色,仅在傍晚或晚上开放。由蜂鸟进行传粉的物种具有一些典型的特征,如具有颜色朝向光谱红色端的花朵,花药和雄蕊从花中突出,形状不呈放射状对称,下唇向下弯曲,产生大量含糖量相对较低的花蜜,如仙人指属、管花柱属(Cleistocactus)和红尾令箭属(Disocactus)中的物种。

停留在仙人掌花朵上的鸟类
蝙蝠授粉在开花植物中相对较少见,但已知仙人掌科种有大约四分之一的属是由蝙蝠进行授粉,比例非常之高。生长在半沙漠地区的柱状仙人掌,是最有可能通过蝙蝠授粉的仙人掌,可能是因为蝙蝠的飞行距离较远,对于分布较稀疏的植物来说,是非常有效的传粉者。由蝙蝠授粉的花朵,通常傍晚和晚上开放,此时蝙蝠较活跃。这些花还有相对暗淡的颜色(通常是白色或绿色);呈放射状对称的形状(通常为管状);一种类似“发霉”的气味;生产大量含糖花蜜等。巨人柱(Carnegiea gigantea)以及一些毛刺柱属(Pilosocereus)和摩天柱属(Pachycereus)的物种均是蝙蝠授粉的记录。

一种南美洲吸食植物花蜜的蝙蝠(Lonchophylla robusta)
栽培
编辑根据仙人掌的原生境不同,可将其分为两类:陆生类型和附生类型。陆生类型产于干旱或半干旱地带,大多数仙人掌都属于该类型。大部分品种为冬季休眠型,在18~20℃以上开始生长,有的12℃以上也缓慢生长,生长期最适合的温度为20~30℃,特别是在昼夜温差大的环境(如白天30~40℃,晚上15~20℃)中,有利于植物积累有机物,更有利于植物生长,但当温度到达38℃以上时,且持续高温闷热特别是延长至晚上时,植物生长缓慢,被迫进入半休眠甚至休眠状态。具有一定的耐寒力,土壤干燥情况下,维持在2~3℃,植物基本不会受害。

南美洲阿根廷城郊的典型仙人掌群落
附生类型产于热带森林中,喜温暖湿润的环境,全年可生长,无休眠现象或休眠不明显,但抗寒和耐高温性较差(13℃以上能正常生长),常攀附生长在大树树干上。虽然附生类型的仙人掌,在自然条件下可能暴露于高光照下,但在人工栽培过程中遇到夏季高温和高光照条件,仍需要适度遮荫,温度不宜上升到32°C以上,仙人掌冬季的最低温度很大程度上取决于其种类。

附生性仙人掌科植物丝苇(Rhipsalis baccifera)
开放基质
仙人掌需要高空气含量的开放基质。当仙人掌在容器中种植时,建议将两部分无土生长介质(水培粘土和分粗浮石或珍珠岩)混合,并添加蚯蚓改善过后的土壤。一般建议包含25-75%的有机物,其余为无机物,如浮石,珍珠岩或砂砾。仙人掌(附生植物除外)喜腐殖质含量低或没有腐殖质的土壤,并建议将粗沙作为生长介质的基础。
人工繁育
人工条件下,仙人掌可以通过种子、扦插或嫁接进行繁育。种子繁殖时将种子播种在潮湿的生长介质中,然后保存在有覆盖的环境中直到种子,以免变干。生长介质不要过分潮湿,否则会导致种子和幼苗腐烂。发芽的适宜温度范围为18~30°C。22°C左右的土温是促进根系生长的最佳温度。在发芽期间,保持较低的光照水平,之后需要提供充足的光照水平以保证其强壮生长。
扦插利用了植物中可以生根的部分。仙人掌会产生易分离或切断的“垫子”“关节”或侧枝。否则,可以进行茎插条,最好是从相对较新的生长中。切割后的扦插组织先进行一段时间的干燥,直到在切割表面上形成愈伤组织。然后可在温度约为22°C的适当生长介质中进行生根。嫁接常用于难以在栽培中生长良好或不能独立生长的物种,例如一些植株呈白色,黄色或红色的无叶绿素植物或某些异常生长的植物(鸡冠状的或畸形的)。砧木需要选择生长强劲并与接穗亲和力较高的植物。
温度光照
仙人掌科植物的栽培温度需要根军原产地来控制,对于大部分原产于热带、亚热带地区的仙人掌科植物 ,一般在 18℃才以上开始生长 ,但温度过高会对植株产生一定的危害。大部分仙人掌科植物生长最适温度为20至30℃,少数种类生长适温在25至35℃,而对于带原产较为寒凉的种类来说,需要将栽培温度维持在15至25℃。绝大多数陆生类型的仙人掌类在生长期间 ,保持较大的昼夜温差 (白天 30 至40℃,晚上15至20℃),对植株生长较为有利。
灌溉浇水
半沙漠仙人掌的灌溉量需要非常注意,频率取决于仙人掌的种植地点、生长介质的性质以及原始栖息地的特点,很难一概而论。仙人掌的死亡原因中,不合时宜的用水占比最多。即使在休眠的冬季,仙人掌也需要水分。在使用硬水灌溉的地方,建议定期换土以避免盐分堆积。在生长季节,仙人掌浇水需要浇透,待变干后再进行浇水。
病虫害防治
病害
真菌、细菌和病毒都会对仙人掌造成危害。在浇水过多时,真菌和细菌易侵染仙人掌。镰刀菌腐病可以通过伤口进入引起腐烂并伴有红紫色霉菌。虫腐病是由双极孢子虫引起的;腐烂病的病菌也会导致仙人掌出现相似的腐烂情况。杀菌剂在对抗这些疾病上的效果非常有限。在仙人掌中发现了几种病毒,包括仙人掌病毒X(CVX),产生少量的外观病状,如绿斑点和马赛克效应(浅色条纹和斑块),尚没有能够有效治疗病毒病的方法。
虫害
害虫主要通过吸食仙人掌汁液对其产生危害,粉蚧主要危害茎和根,鳞虫主要危害茎,棉红蜘蛛,数量虽小但可大量出现,它们在自己周围构建一个细网,并通过吸吮汁液来严重标记仙人掌;蓟马主要危害花朵。其中一些害虫,对许多杀虫剂具有抗药性。眼蕈蚊和蕈蚊的幼虫会食用仙人掌的根,蛞蝓和蜗牛食用仙人掌。
保护
编辑仙人掌科植物,在野外面临的主要威胁主要由于其生长发育、人类放牧和过度采集等产生的问题。在墨西哥基马潘附近建造的大坝,导致金琥(Echinocactus grusonii)的大部分自然栖息地遭到破坏;城市发展和高速公路导致墨西哥、新墨西哥州和亚利桑那州部分地区的仙人掌栖息地,如索诺兰沙漠等被破坏;发展农业也会影响仙人掌科植物的生长,墨西哥的干旱平原被用来种植玉米后,黑牡丹(Ariocarpus kotschoubeyanus)的数量受到影响,智利山谷的山坡上种植藤曼后的龙爪球属(Copiapoa)和壶花柱属(Eulychnia)植物的数量受到影响。在许多地区,放牧引入的动物(如山羊)对仙人掌的种群造成了严重的损害。过度采集仙人掌进行出售对一些物种造成了极大的影响。由于过度采挖,导致墨西哥米基瓦纳附近的银牡丹(Pelecyphora strobiliformis)几乎消失。

通往美国新墨西哥州陶斯的高速公路
保护名录
仙人掌科的所有种均被列入《濒危野生动植物种国际贸易公约》。若无出口许可证,大多数仙人掌标本在国际贸易中都是非法的。也存在一些例外,比如“归化或人工繁殖的植物”。一些仙人掌种类,如岩牡丹属和圆盘玉属(Discocactus)中的所有物种,因为其濒危程度均被列入更严格的附录I中,这些物种只能出于非商业目的在双方均含有进口或出口许可证的国家之间流转。除被列入附录Ⅰ的物种和木麒麟属所有种、麒麟掌属所有种以及船夫掌属(Quiabentia)所有种外,仙人掌科所有种均被列入《濒危野生动植物种国际贸易公约》(CITES)(2019)的附录II中。
保护措施
仙人掌的保护,有就地保护和迁地保护两种方式。就地保护是指通过实施法律保护和在原自然生境建立保护区来保护植物,如美国建立的德克萨斯州的大弯国家公园、加利福尼亚州约书亚树国家公园和亚利桑那州的萨瓜罗国家公园,墨西哥建立的索诺拉州皮纳卡特国家公园,智利建立的的潘德阿苏卡尔国家公园。迁地保护则是将植物和种子保存在其自然生境之外,如人掌和其他多肉植物的种子被长期储存在美国亚利桑那州的沙漠植物园中。
用途
编辑栽培食用
大部分肉质的仙人掌果实都是可食用的。巨人柱(Carnegiea gigantea)的果实是墨西哥西北部和美国西南部(包括索诺拉沙漠)的土著部落中重要的植物资源。人们通过煮沸制糖浆和干燥两种方式来保存其果实,制得的糖浆可以用来发酵生产酒精饮料。新绿柱属(Stenocereus)植物的果实也是北美地区的重要食物来源。在更热带的南部地区,攀援植物量天尺(Hylocereus undatus)的果实可食用,在亚洲广泛种植,商品名为火龙果。除了新绿柱属外,鹿角柱属(Echinocereus)、强刺球属(Ferocactus)、乳突球属(Mammillaria)、龙神柱属(Myrtillocactus)、摩天柱属(Pachycereus)、块根柱属(Peniocereus)和蛇鞭柱属(Selenicereus)中某些物种的果实也可食用。
梨果仙人掌(Opuntia ficus-indica)长期以来都是当地重要的食物来源,其原始种来自墨西哥中部,北美南部的土著在此基础上培育出了一系列的栽培品种。梨果仙人掌的果实和叶枕均可食用,并命名为tuna和nopal。2007年墨西哥的nopal产业价值可达1.5亿美元。此外除了美洲,梨果仙人掌也是西西里岛、阿尔及利亚和其他北非国家的重要经济作物。仙人掌属的其他物种的果实也可食用,通常也冠以相同的名称(tuna)。
建筑材料
仙人掌还被用作建筑材料。活仙人掌围栏被用作建筑物周围的路障,以防止人们闯入或用来圈养动物。仙人掌如秘鲁天伦柱(Cereus repandus)等植物的木质部分,可用于建筑物和家具中,如墨西哥塞里人会使用巨人柱(Carnegiea gigantea)来建造篱笆房屋的框架。一些仙人掌的细刺和毛状体还可用作填充枕头和编织过程中的纤维来源。
园艺观赏
仙人掌首次从美洲引种就是因为其观赏价值。1800年代初,欧洲的爱好者拥有大量仙人掌收藏品,稀有品种还以非常高的价格出售。仙人掌的供应商除了自己种植外,还雇用收集者从野外获取植物。
改善生态
许多仙人掌可以忍受长时间的干燥环境,在干旱缺水或水土流失严重的区域,可利用仙人掌这类抗旱植物来促进改善生态结构,促进植被恢复和打造旱地景观。
其他用途
许多种类的仙人掌已被证明含有某些精神类活性成分,这些化合物可以通过对大脑的影响引起情绪、感觉和认知的变化。在美洲的土著部落中乌羽玉(Lophophora williamsii)和毛花柱(Echinopsis pachanoi)均有悠久的使用历史,两者都含有麦司卡林(仙人掌毒碱)。仙人掌在烧掉刺后还可作为食物或动物的饲料,一些仙人掌还可做草药。仙人掌上生活着一种名为胭脂虫的介壳虫,可提取红色染料,长期为美洲和北美人民所用,直到人工合成的煤焦油类染料的出现,其需求才下降。随着对天然染料需求的增加,商业生产在逐渐增加。
危害
编辑在澳大利亚,仙人掌属植物,特别是缩刺仙人掌(Opuntia stricta),于19世纪被引入,用作天然农用栅栏,并尝试建立胭脂虫产业。这些外来种迅速成为当地的主要杂草,但现在因为生物防治(如一种螟蛾Cactoblastis cactum)的应用而得到控制。然而,澳大利亚仙人掌物种的杂草危害仍存在,2012年四月,澳大利亚杂草委员会宣布除梨果仙人掌以外的所有仙人掌均为具有国家意义的杂草。此外阿拉伯半岛的仙人掌种群仍在不断增加。其中既有栽培的,也有逃逸到野外的。
在中国,仙人掌属(Opuntia)共有 3 种入侵植物, 分别是仙人掌(Opuntia dillenii)、 梨果仙人掌(Opuntia ficus-indica)和 单刺仙人掌(Opuntia monacantha),中国学者李振宇和解焱(2002)报道仙人掌在广东、 香港、澳门、广西南部、海南沿海地区及南海诸岛逸为野生,随后仙人掌被多次报道在中国多个省份入侵。
文化
编辑仙人掌被人类最早的种植时间没有确定的结论。北美洲南部墨西哥的一支印第安人阿兹特克人,将仙人掌属用于了多种场景中,他们的首都是特诺奇蒂特兰(现墨西哥城),其名字起源的一种解释就是,涵义是纳瓦特语的nichtli(一种仙人掌的果实)。墨西哥国徽上,有鹰栖息在仙人掌上,抓握着一条蛇的图案,源于古都特诺奇蒂特兰建国神话。阿兹特克人象征性地将成熟的红色仙人掌果实与人类的心脏联系在一起,向太阳神献上人类的心可以确保太阳继续运转,就像水果可以解渴一样。
参考资料
编辑展开[1]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 611.
[2]Species.物种2000中国节点. [2023-02-23].
[3]仙人掌科.多识植物百科. [2023-02-23].
[4]仙人掌科 (xiān rén zhǎng kē).植物智. [2023-12-20].
[5]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 611.
[6]仙人掌科 (xiān rén zhǎng kē).植物智. [2023-02-23].
[7]Shetty A A, Rana M K, Preetham S P. Cactus: a medicinal food[J]. Journal of food science and technology, 2012: 530-536.
[8]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 566.
[9]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[10]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[11]Raven J A, Edwards D. Roots: evolutionary origins and biogeochemical significance[J]. Journal of experimental botany, 2001, 52(suppl_1): 381-401.
[12]Sharkey T D. Estimating the rate of photorespiration in leaves[J]. Physiologia Plantarum, 1988, 73(1): 147-152.
[13]Edwards E J, Nyffeler R, Donoghue M J. Basal cactus phylogeny: implications of Pereskia (Cactaceae) paraphyly for the transition to the cactus life form[J]. American Journal of Botany, 2005, 92(7): 1177-1188.
[14]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 51.
[15]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 61.
[16]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 61.
[17]张忠和, 石雷, 徐珑峰, 王自力. 世界胭脂虫的研究和利用概况[J]. 林业科学研究, 2002, (06): 719-726.
[18]傅佑丽. 仙人掌资源的价值分析及开发利用[J]. 国土与自然资源研究, 2007: 93-94. DOI:10.16202/j.cnki.tnrs.2007.03.005.
[19]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 45-49.
[20]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[21]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[22]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[23]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[24]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[25]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 398.
[26]Mauseth J D. Tiny but complex foliage leaves occur in many “leafless” cacti (Cactaceae)[J]. International Journal of Plant Sciences, 2007, 168(6): 845-853.
[27]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[28]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[29]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 566.
[30]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 347.
[31]Edwards E J, Donoghue M J. Pereskia and the origin of the cactus life-form[J]. The American Naturalist, 2006, 167(6): 777-793.
[32]Edwards E J, Donoghue M J. Pereskia and the origin of the cactus life-form[J]. The American Naturalist, 2006, 167(6): 777-793.
[33]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 572.
[34]Mauseth J D. Tiny but complex foliage leaves occur in many “leafless” cacti (Cactaceae)[J]. International Journal of Plant Sciences, 2007, 168(6): 845-853.
[35]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[36]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[37]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[38]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[39]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 174.
[40]Keeley J E, Rundel P W. Evolution of CAM and C4 carbon-concentrating mechanisms[J]. International journal of plant sciences, 2003, 164(S3): S55-S77.
[41]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 39.
[42]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 611.
[43]Cota-Sánchez J H, Bomfim-Patrício M C. Seed morphology, polyploidy and the evolutionary history of the epiphytic cactus Rhipsalis baccifera (Cactaceae)[J]. Polibotánica, 2010, (29): 107-129.
[44]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 399.
[45]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 174.
[46]Eawards E J, Nyffeler R, Donoghue M J. Basal cactus phylogeny: implications of Pereskia (Cactaceae) paraphyly for the transition to the cactus life form[J]. American Journal of Botany, 2005, 92(7): 1177-1188.
[47]Rowley, Gordon D. A History of Succulent Plants[M]. Calif: Strawberry Press, 1997: 43.
[48]Raven J A, Edwards D. Roots: evolutionary origins and biogeochemical significance[J]. Journal of experimental botany, 2001, 52(suppl_1): 381-401.
[49]Sharkey T D. Estimating the rate of photorespiration in leaves[J]. Physiologia Plantarum, 1988, 73(1): 147-152.
[50]李振宇, 解焱. 中国外来入侵种[M]. 北京: 中国林业出版社, 2002: 130-132.
[51]李惠茹. 我国仙人掌属植物的入侵性研究[J]. 园艺与种苗, 2018: 24-27. DOI:10.16530.
[52]Johnson A T, Smith H A. Plant Names Simplified : Their Pronunciation Derivation & Meaning[M]. 1972: 19.
[53]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 93-98.
[54]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 93-98.
[55]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 99.
[56]Bárcenas R T, Yesson C, Hawkins J A. Molecular systematics of the Cactaceae[J]. Cladistics, 2011, 27(5): 470-489.
[57]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 99.
[58]Mayta L, Molinari-Novoa E A. L’intégration Du Genre Leuenbergeria Lodé Dans Sa Propre Sous-Famille, Leuenbergerioideae Mayta & Mol[J]. Nov., 2015: 6-7.
[59]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 99.
[60]Edwards E J, Donoghue M J. Pereskia and the origin of the cactus life-form[J]. The American Naturalist, 2006, 167(6): 777-793.
[61]dwards E J, Nyffeler R, Donoghue M J. Basal cactus phylogeny: implications of Pereskia (Cactaceae) paraphyly for the transition to the cactus life form[J]. American Journal of Botany, 2005, 92(7): 1177-1188.
[62]Bárcenas R T, Yesson C, Hawkins J A. Molecular systematics of the Cactaceae[J]. Cladistics, 2011, 27(5): 470-489.
[63]Lodé J. Leuenbergeria, a new genus in Cactaceae[J]. Cact. Advent. Int, 2013: 25-27.
[64]ASAI I, MIYATA K. An Emendation of Rhodocactus, a Genus Segregated from Pereskia (Cactaceae)[J]. J. Jpn. Bot, 2016: 7-12.
[65]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 99.
[66]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 399.
[67]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 485.
[68]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 398.
[69]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 99.
[70]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 99.
[71]Arakaki M, Christin P A, Nyffeler R, et al. Contemporaneous and recent radiations of the world's major succulent plant lineages[J]. Proceedings of the National Academy of Sciences, 2011, 108(20): 8379-8384.
[72]Rowley, Gordon D. A History of Succulent Plants[M]. Calif: Strawberry Press, 1997: 43.
[73]Edwards E J, Nyffeler R, Donoghue M J. Basal cactus phylogeny: implications of Pereskia (Cactaceae) paraphyly for the transition to the cactus life form[J]. American Journal of Botany, 2005, 92(7): 1177-1188.
[74]Arakaki M, Christin P A, Nyffeler R, et al. Contemporaneous and recent radiations of the world's major succulent plant lineages[J]. Proceedings of the National Academy of Sciences, 2011, 108(20): 8379-8384.
[75]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 174.
[76]Johnson A T, Smith H A, Stockdale A P. Plant Names Simplified: Their Pronunciation, Derivation & Meaning[M]. Their Pronunciation, 2019: 26.
[77]Arakaki M, Christin P A, Nyffeler R, et al. Contemporaneous and recent radiations of the world's major succulent plant lineages[J]. Proceedings of the National Academy of Sciences, 2011, 108(20): 8379-8384.
[78]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[79]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[80]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[81]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 15.
[82]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 33.
[83]Hartmann S, Nason J D, Bhattacharya D. Phylogenetic origins of Lophocereus (Cactaceae) and the senita cactus–senita moth pollination mutualism[J]. American Journal of Botany, 2002, 89(7): 1085-1092.
[84]Fenster C B, Armbruster W S, Wilson P, et al. Pollination syndromes and floral specialization[J]. Annu. Rev. Ecol. Evol. Syst., 2004: 375-403.
[85]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 33.
[86]McMillan A J S, Horobin J F. Christmas Cacti : The genus Schlumbergera and its hybrids[M]. 1995: 49.
[87]Fleming T H, Geiselman C, Kress W J. The evolution of bat pollination: a phylogenetic perspective[J]. Annals of botany, 2009, 104(6): 1017-1043.
[88]罗婵娟. 仙人掌科多肉植物在福州园林应用中的研究[J]. 福建建材, 2021, (06): 63-66+111.
[89]Brown, Roger. Cultivation of Cacti[M]. 2001: 85.
[90]Hewitt, Terry. The Complete Book of Cacti & Succulents[M]. London: Covent Garden Books, 1993: 150-151.
[91]Anderson, Miles. Cacti and Succulents : Illustrated Encyclopedia[M]. 1999: 217.
[92]Anderson, Miles. Cacti and Succulents : Illustrated Encyclopedia[M]. 1999: 217.
[93]Hewitt, Terry. The Complete Book of Cacti & Succulents[M]. London: Covent Garden Books, 1993: 147.
[94]Innes, Clive & Wall, Bill. Cacti, Succulents and Bromeliads[M]. London: Cassell for the Royal Horticultural Society, 1995: 23.
[95]Keen, Bill. Cacti and Succulents : step-by-step to growing success[M]. 1990: 27.
[96]Hecht, Hans. Cacti & Succulents[M]. New York: Sterling, 1994: 140.
[97]Brown, Roger. Cultivation of Cacti[M]. 2001: 85.
[98]Innes, Clive & Wall, Bill. Cacti, Succulents and Bromeliads[M]. London: Cassell for the Royal Horticultural Society, 1995: 27-29.
[99]Brown, Roger. Cultivation of Cacti[M]. 2001: 85.
[100]Innes, Clive & Wall, Bill. Cacti, Succulents and Bromeliads[M]. London: Cassell for the Royal Horticultural Society, 1995: 27-29.
[101]nnes, Clive & Wall, Bill. Cacti, Succulents and Bromeliads[M]. London: Cassell for the Royal Horticultural Society, 1995: 27-29.
[102]鲍海鸥, 陈波红. 仙人掌科植物的栽培技术[J]. 江西园艺, 2000, (2): 31-33.
[103]Pilbeam, John. Cacti for the Connoisseur[M]. London: Batsford, 1987: 10.
[104]Brown, Roger. Cultivation of Cacti[M]. 2001: 85.
[105]Hewitt, Terry. The Complete Book of Cacti & Succulents[M]. London: Covent Garden Books, 1993: 150-151.
[106]Brown, Roger. Cultivation of Cacti[M]. 1995: 85.
[107]Bipolaris cactivora (Petr.) Alcorn.GBIF. [2023-02-27].
[108]Hecht, Hans. Cacti & Succulents[M]. New York: Sterling, 1994: 152.
[109]Duarte L M L, Alexandre M A V, Rivas E B, et al. Potexvirus diversity in Cactaceae from Sao Paulo state in Brazil[J]. Journal of Plant Pathology, 2008: 545-551.
[110]Hecht, Hans. Cacti & Succulents[M]. New York: Sterling, 1994: 152.
[111]Innes, Clive & Wall, Bill. Cacti, Succulents and Bromeliads[M]. London: Cassell for the Royal Horticultural Society, 1995: 31-32.
[112]Innes, Clive & Wall, Bill. Cacti, Succulents and Bromeliads[M]. London: Cassell for the Royal Horticultural Society, 1995: 31-32.
[113]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 73.
[114]2019年CITES附录中文版.中华人民共和国濒危物种科学委员会. [2023-02-27].
[115]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 73.
[116]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 51.
[117]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 51.
[118]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 51.
[119]Griffith M P. The origins of an important cactus crop, Opuntia ficus‐indica (Cactaceae): new molecular evidence[J]. American Journal of Botany, 2004, 91(11): 1915-1921.
[120]nderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 51.
[121]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 61.
[122]Keen, Bill. Cacti and Succulents : step-by-step to growing success[M]. 1990: 15.
[123]傅佑丽. 仙人掌资源的价值分析及开发利用[J]. 国土与自然资源研究, 2007: 93-94. DOI:10.16202/j.cnki.tnrs.2007.03.005.
[124]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 45-49.
[125]Shetty A A, Rana M K, Preetham S P. Cactus: a medicinal food[J]. Journal of food science and technology, 2012: 530-536.
[126]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 61.
[127]张忠和, 石雷, 徐珑峰, 王自力. 世界胭脂虫的研究和利用概况[J]. 林业科学研究, 2002, (06): 719-726.
[128]Weed Identification.Australian Weeds Committee. [2023-02-27].
[129]Al-Robai S A, Howladar S M, Mohamed H A, et al. ) Backeb,(Cactaceae): a new generic alien record in the flora of Saudi Arabia[J]. Journal of Asia-Pacific Biodiversity, 2018: 320-323.
[130]El-Hawagry M S, Al Dhafer H M. , a new record of the subfamily Phthiriinae (Bombyliidae, Diptera) from Saudi Arabia[J]. Zootaxa, 3872, (4): 387-392.
[131]李惠茹. 我国仙人掌属植物的入侵性研究[J]. 园艺与种苗, 2018: 24-27. DOI:10.16530.
[132]Anderson, Edward F. The Cactus Family[M]. Oregon: Timber Press, 2001: 485.
[133]Arakaki M, Christin P A, Nyffeler R, et al. Contemporaneous and recent radiations of the world's major succulent plant lineages[J]. Proceedings of the National Academy of Sciences, 2011, 108(20): 8379-8384.
[134]Johnson A T, Smith H A, Stockdale A P. Plant Names Simplified: Their Pronunciation, Derivation & Meaning[M]. Their Pronunciation, 2019: 26.
该页面最新编辑时间为 2024年4月2日
百科词条作者:小小编,如若转载,请注明出处:https://glopedia.cn/71729/