
环节动物门(学名:Annelida)是动物界下的一门,环节动物演化出的真体腔和身体分节现象,使其在动物演化上占重要位置。截至2024年,在国际综合分类学信息系统(ITIS)的分类标准中,节肢动物门共有2纲、17目、152科、1438属、9466种和800个亚种。
起源
编辑一般认为环节动物起源于扁形动物门(Platyhelminthes)的涡虫纲(Turbellaria),因为环节动物和涡虫的胚胎发育都经过螺旋型卵裂;环节动物多毛纲的担轮幼虫和扁形动物的牟勒氏幼虫很相似;有些环节动物还保留了具有管细胞的原肾管,与扁形动物的焰细胞原肾管在本质上相同。另一学说认为环节动物来自一种现已不存在的假设的担轮动物,该物种与现存的轮虫动物门的球轮虫很相似。化石纪录表明,所有的环节动物都起源于前寒武纪的海洋中(分子测年法表明环节动物最早出现于埃迪卡拉纪),直到奥陶纪和泥盆纪后期才有陆生环节动物的出现,说明陆生环节动物是水生种类的后代。
现存的三个纲中,多毛类比较原始,具担轮幼虫,自寒武纪以来,多毛类动物开始逐渐向海洋、淡水和陆地环境中扩散,是海洋泥沙底质、沉积岩乃至远洋海底的主要组成部分。寡毛类是较早从多毛类分出的一支,适应陆地穴居生活,头部感官退化,无疣足而有刚毛。蛭类和寡毛类亲缘关系较近,可能是由原始寡毛类演化而来。
也有认为寡毛类是较原始的一纲,据推测,原始环节动物同律分节,体腔分隔,有成对的上皮刚毛,由口前叶和围口节构成头部,与现代寡毛类很类似;多毛类的疣足和发达的头部是在称为现代多毛类的过程中发展的。


分类
编辑截至2024年,在国际综合分类学信息系统(ITIS)的分类标准中,环节动物门共有2纲、17目、152科、1438属。已知种类约有17000余种或22000余种。按照新的分类系统,本门分环带纲(Clitellata)和多毛纲(Polychaeta)2纲,传统分类系统将环带纲分为寡毛纲(Oligochaeta)和蛭纲(Hirudinea)2纲。截止至2023年,按ITIS分类标准,环带纲和多毛纲两纲分类如下:
| 环带纲 | 多毛纲 |
| Tubificata亚纲 | 螠虫亚纲(Echiura) |
| Randiellata亚纲 | 游走亚纲(Errantia) |
| LumbrICUlata亚纲 | 隐居亚纲(Sedentaria) |
传统分类系统中,多毛纲下的螠虫亚纲仍作为螠虫动物门单独分类,后按此分类系统介绍,此分类系统下,多毛纲、寡毛纲和蛭纲三纲分类如下:
| 多毛纲 | 寡毛纲 | 蛭纲 |
| 蠕形亚纲(Scolecida) | 水蚓目(Limnicolae) | 棘蛭目(Acanthobdellida) |
| 触须亚纲(Palpata) | 陆蚓目(Terricolae) | 鳃蛭目(Branchiobdellida) |
| 水蛭目(Hirudinida) |
其中多毛纲包括6000多种已知物种,寡毛纲有约3250种已知物种,蛭纲有约300种。
形态特征
编辑环节动物的身体由许多形态相似的体节构成,是为分节现象,每个体节之间在体内以隔膜分隔,体表相应形成节间沟,为体节的分界,虫体最前端的口前叶和最后端的尾节在外形上酷似体节,但一般不认为是体节,虫体内部结构也多是分节存在,器官如附肢、体腔、肌肉、肾管和生殖腺等通常在每个体节内重复出现,这种各体节在形态和功能上基本相似的分节现象称为同律分节,环节动物一般为两侧对称,具有真体腔,体长从几毫米到3米不等。
环节动物发展出了原始的附肢,及疣足和刚毛,海生种类一般每一体节具有一对疣足,疣足是由体壁向外突出的扁平叶状结构,可分为背叶和腹叶,背叶的背侧和腹叶的腹侧有一指状的背须和腹须,有些种类的背须特化为疣足鳃或鳞片等,背叶和腹叶的边缘各有一束刚毛;陆地和淡水生活的水蛭和蚯蚓等没有疣足,但蚯蚓有刚毛,自上皮内陷的刚毛囊伸出,是由囊底部的一个成刚毛细胞分泌的几丁质构成的,疣足和刚毛的出现,增强了动物的运动能力。

异色菏沙蚕(Hediste diversicolor)
体壁和真体腔
环节动物在进化上的重要特征是具有端细胞法形成的真体腔,又称次生体腔。环节动物每个体节内有两侧成对的体腔,每个相邻体节的体腔由隔膜分开,每一体节的左右体腔以背、腹系膜分开,具疣足的动物,其体腔会伸入疣足种。真体腔内充满体腔液,其内含有体腔细胞,有内部防伪功能;有的体腔细胞含有血红蛋白,也有气体运输功能。环节动物的体腔液具有强有力的流体静力骨骼作用,在运动时可迅速将肌肉收缩的力传至身体各处,增强了动物体的运动机能。
真体腔和体节的出现除提高运动的灵活性和有效性外,还促进了动物结构和机能多方面的复杂化,如消化管壁出现的肌肉层,增强了蠕动,提高了消化机能,同时真体腔的出现还促进了循环、排泄和神经系统的发展,使动物体结构更加复杂,各种机能趋于完善。
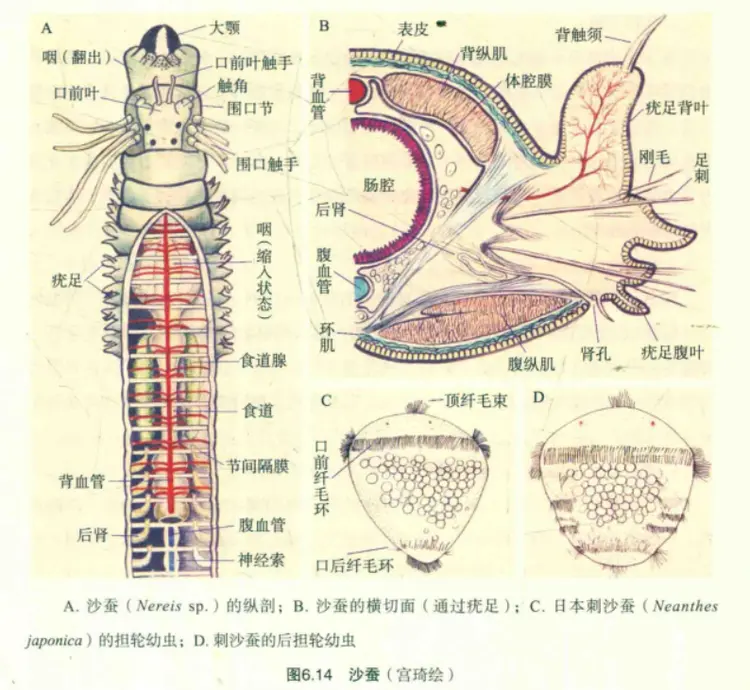
沙蚕体腔手绘图——环节动物的体壁由外及内包括角质层、表皮层、不明显的基膜/环肌层、纵肌层和体腔膜
环节动物的体壁由外及内包括角质层、表皮层、不明显的基膜/环肌层、纵肌层和体腔膜。
循环系统
环节动物循环系统的形成与真体腔发生密切相关,典型的循环系统是闭管式循环系统。由纵行血管和环行血管及其分支血管以及与各血管相连的微血管网组成,血液始终在管内流动,不流入组织间的空隙中,故称闭管式循环。血液循环有一定方向,流速较恒定,一般环节动物(除最原始种类外)血浆中含有血红蛋白、蚯蚓血蓝蛋白和血绿蛋白3种呼吸色素。有一些环节动物(蛭类)的真体腔被结缔组织填充,形成了不同的腔隙,血液在其中流动,实际血液为血体腔液。
消化系统
消化系统简单,一般由口、咽、食道、胃、肠、直肠和肛门组成,蚯蚓等陆生种类在食道后特化出嗉囊和砂囊,蛭类等寄生种类消化系统则由口、口腔、咽、食道、嗉囊、肠和肛门组成,环节动物的消化管外附有肌肉层,不随体节分节。肛门位于体后端,管栖的多毛类动物通常会在管中留出粪便通道,使沉积在管底的粪便等沿通道向管前端排出。
呼吸器官
环节动物无完整的呼吸系统,通常通过体表和外界进行气体交换,但有些多毛类具有特化的鳃,或由疣足表面密布的血管充当呼吸器官,水生寡毛类的直肠也有气体交换的功能。
排泄系统
环节动物的排泄系统为后肾管型,按体节排列,每节一对或多个,典型后肾管为一条盘曲迂回的管子,一端开口于前一体节的体腔称为肾口,具有带纤毛的漏斗,另一端开口于本体节腹侧体表,称为肾孔,这样的肾管常称为大肾管,体腔液从肾口大量进入肾管腔种,通过其上密布的血管网有选择地将体腔液重吸收回体腔中;有些种类后肾管特化为小肾管,无肾口,肾孔开口于体壁或开口于消化道;原始种类在每个体节上除一对肾管外还有一对由中胚层形成的体腔管,一端开口于体腔,另一端开口于体表;有的种类原肾管或后肾管与体腔管结合,分别称为混合原肾管和混合后肾管,多数后肾管与体腔管高度融合为后肾管。
神经系统
环节动物的神经系统为索式神经系统,每一体节都有一对神经节,体前端背侧有一对咽上神经节或称脑神经节,左右由一对围咽神经与一对已愈合的咽下神经节相连,咽下神经节是腹神经索的第一个神经节,由此向后的腹神经索纵贯全身(腹神经索由两条纵行的副神经合并而成);脑神经节分化为三个区域,前脑中脑和后脑,前脑支配口前叶的感觉器,中脑支配眼和口前叶的触手或触须,并分出交感神经到消化管,可控制吻、咽的活动,围咽神经就是由前脑和中脑发出的,后脑支配化学感受器。
感觉器官
环节动物的感觉器官有触须、味蕾、平衡囊、感光细胞以及眼点,多毛纲的一些种类在口前叶后端两侧有一对纤毛窝,称为项器,是化学感受器,有嗅觉功能。游泳生活的多毛纲动物眼常分布在头部,偶尔分布在鳃和身体两侧,而固着生活的多毛类眼常退化,依靠触须进行感知食物等活动;寡毛纲动物的眼散布在身体上;蛭纲动物的眼成对位于头部前端。
分布栖息
编辑分布范围
环节动物门物种在世界各地均有分布,已发现的物种主要集中在欧洲东部地区、北美洲南部沿海和亚洲东南部。其中多毛纲动物广泛分布于各大海区;寡毛纲动物则在除沙漠外的陆地均有分布。
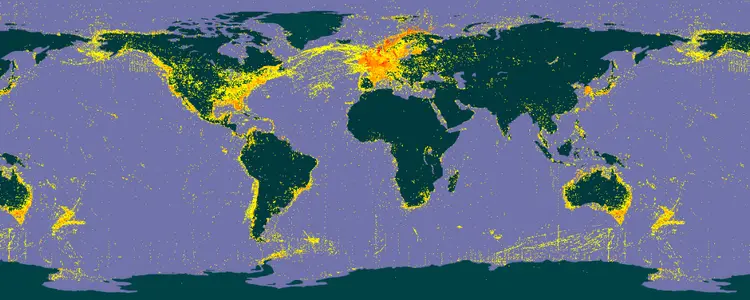
已发现的环节动物在世界范围内的分布
栖息环境
环节动物门栖息于陆地、淡水、海洋等各种生境,少数寄生生活:
多毛纲中的许多种类都可以自由生活在浅海海底表面、石块或贝壳下、珊瑚礁及海藻等植物表面,例如沙蚕科(Nereidae)等,此类多毛纲动物口前叶具触手、眼等感觉器官,疣足发达,躯干部体节相似,一般善于运动;一些种类在大洋中营浮游生活,例如浮蚕科(Tomopteridae)等,身体通常透明;一些种类钻穴生活,如吻沙蚕科(Glyceridae)等,在海底泥沙中钻穴移动时可同时分泌黏液,形成一黏液环绕的走道并在其中生活,通常感觉器官不发达;一些种类营管居生活,以自身分泌的黏液黏着外界的有机物或无机物形成各种形态和质地的管道并将其作为巢穴,如巢沙蚕(Diopetra);一些种类与其他生物有共生或寄生关系,许多管居或穴居的多毛类动物,其管道或穴道内共生有甲壳类、软体动物或其他多毛类动物,而如鱼沙蚕则寄生在海鳝体表。
寡毛纲大多在于潮湿、富含有机质的中性土壤中营穴居生活,除沙漠地区外,任何土壤中都有分布,陆生种类约占此纲的80%;也有少部分为淡水生,极少数生活在海水或微咸水中,一般水生种类体型较小、结构简化;也有寄生种类。
蛭纲大多数生活在淡水中,少数生活在海洋中,还有部分栖息在森林或草丛中,多数营暂时性体外寄生生活,也有永久性寄生的种类。
| 潮间带的沙蚕科的旗须沙蚕(Nereis vexillosa)
|
| 浮游生活的浮蚕科的生殖浮蚕(Tomopteris helgolandica)
|
| 钻穴生活的吻沙蚕科的加州半足沙蚕(HemiPodia californiensis)
|
| 铜色巢沙蚕(Diopatra cuprea)利用贝壳等黏合形成的管状巢穴
|




生活习性
编辑环节动物不同纲生活习性差异较大。
摄食及相应结构
多毛类动物栖息方式的多样化伴随着食性的多样化,腐食性、植食性、捕食性、滤食性乃至渗透营养性生物均有发现。根据体型的不同,小体型的多毛纲动物会以捕食或滤食或二者兼备的方式以浮游生物为食,或者以底上或底下的底栖生物为食,还有的种类连海底的泥沙一同吞下,将其中的有机物消化后排出;而大体型的多毛纲动物则分为肉食性、腐食性、寄生性和植食性,其中植食性多毛纲动物根据栖息地不同,有三种不同的食物来源,一是光合自养的小型生物,二是巨藻类,三是被子植物;多毛纲动物在不同的生命阶段会周期性更换食物来源,有些种类则在同一生命阶段同时有多种食物来源;除此之外,多毛纲还有一种营渗透营养的物种,巨型管虫(Riftia pachyptila)是世界上已知最大的渗透性营养生物,无口以及消化系统。

巨型管虫
与食性相适应,小型多毛纲动物咽部通常缺乏肌肉,取而代之的是一些种类在咽部会有纤毛着生;大体型的多毛纲动物通常咽部肌肉发达;而渗透营养的巨型管虫则咽部退化消失,体内多环形褶皱,增大体内化能合成生物的附着面积。
陆生的寡毛纲动物通常为食腐动物,以腐烂的有机物、植物叶茎碎片为食,咽部有分泌粘液和蛋白酶的单细胞咽腺,可润湿食物和初步消化;咽壁厚肌肉质,可将咽腔扩大,将食物吞下,咽后的食道壁有钙腺,能分泌钙离子到消化管,中和土壤中的酸性物质。
水生寡毛纲动物体型较小,但食性与陆生种类相似,以水底淤泥为食料。
蛭纲动物少数为捕食性,取食小型的蠕虫、螺类及昆虫的幼虫,75%的种类营半寄生性吸血生活,其中较原始的种类吸食各种无脊椎动物的血液,较高等的种类吸食各种脊椎动物的血液。
蛭类体前端和后端各有一吸盘,有吸附功能,口位于前吸盘中央,口腔后为肌肉质的咽,咽壁肌肉发达,便于抽吸血液。一些种类(如吻蛭目Rhynchobdellida)有一个肌肉质,可向外翻出的吻,取食时将吻刺入宿主吸食血液,吻腔内有单细胞腺可分泌蛭素防止血液凝固,舌蛭科(Glossiphoniidae)下的一些物种还可以用吻将猎物包裹后吞入;另一些(如颚蛭目 Gnathobdellida)种类无吻,口腔内有三个呈三角形排列的颚,用以造成伤口,咽壁周围有单细胞分泌腺,用以分泌蛭素。捕食性的蛭类胃为一简单的直管,吸血种类的胃变成了有1~11对侧盲囊的嗉囊,其中最后一对直达身体后端,用途是储存血液。
再生
环节动物通常可以利用一部分组织将身体完全再生,其中多毛类和低等寡毛类动物的再生能力强于高等寡毛类,而蛭类则再生能力较弱。大部分多毛类和寡毛类动物前端可以再生身体缺失的后部,再生后的体节数不会超过受伤前的体节数。
再生的过程一般可概括为:伤口封闭、形成芽体、中胚层再生细胞的迁移,随即是对缺失部分的修复。
再生能力也给环节动物带来了较长的寿命:许多多毛类动物在繁殖一次后就会死亡的情况下,最长寿的物种寿命甚至可以达到三年(围沙蚕属 Perinereis);一种水栖寡毛类在实验条件下无性繁殖了130代;一些蚯蚓被认为寿命可以达到10年。但截至2024年,人们对蛭类动物的寿命尚无较准确的了解。
发光现象
一些种类的多毛纲动物(如裂虫属 Syllis、毛翼虫属 Chaetopterus 等)具有自发光现象,不同种类的发光方式各有不同:Polynoe 的光来自于细胞内;一些海鳞虫属(Halosydna)动物皮下具有特殊的光电管,在受到刺激时会发光;齿裂虫属(Odontosyllis)动物发出的光与性成熟和群游现象(受月球周期的影响)有关,雌性会发出更明亮的光,以吸引发光的雄性,释放配子后,雌性的发光现象会减弱;在毛翼虫科(Chaetopterida)中,发光现象通常是某些体节和触角排出的发光分泌物造成的,这种分泌受神经的控制,如毛翼虫属动物可以通过刺激腹侧神经使附肢发光。
寡毛纲也有发光的现象,当受到刺激时,一些蚯蚓会从口腔、肛门、背孔或排泄孔产生发光黏液,发光黏液可能是由其体内共生的细菌产生的。
蛭类没有已知种类出现过发现发光现象。
生长繁殖
编辑个体发育
环节动物的受精卵经螺旋卵裂、定型发育,通过内陷或外包或两者结合形成原肠胚,此后陆生和淡水种类直接发育为成虫,海产种类胚胎迅速发育为担轮幼虫,一般担轮幼虫形似陀螺,体分为三部分:口前纤毛区,包括口前纤毛环、口和感觉板;口后纤毛区,包括口后纤毛环和肛门区;生长带区,包括口前纤毛环和口后纤毛环之间的区域。变态发育时,口前纤毛区发育为成体的口前叶及其触手等感觉器,口后纤毛区发育为尾节,生长带区细胞不断分裂增殖、分化成躯干的所有体节和其体腔。
真体腔的形成
在胚胎或幼虫期,节生长区成对的端细胞分裂增殖形成左右两中胚层带,继续发育,左右中胚层带内逐渐充以液体,并分节裂开,形成每节一对体腔,此时真体腔位于中胚层之间,是由中胚层裂开形成的体腔,故又名裂体腔;每个体腔继续发育扩大,其外侧的中胚层附在外胚层的内面,分化成肌肉层和体腔膜,与肠上皮构成肠壁,在每个体腔的前后按体节由体腔膜形成双层的隔膜,在背、腹侧与消化道结合形成背系膜和腹系膜。故环节动物的身体生长是从后端连续增加体节的结果,即最年轻的体节位于尾节之前,最老的体节是位于口前叶之后的第一个体节,这种生长模式称为端细胞生长。
生殖
多毛纲
多毛类大多数为雌雄异体,无固定的生殖腺和生殖导管,在生殖季节,生殖腺来自体腔上皮(消化道周围的中胚层),精子由肾管排出;卵成熟后由体背侧鳞式开口排出,在海水中受精。少数种类(通常是固着生活种类)能行无性生殖,主要进行出芽生殖或分裂生殖,如裂虫和自裂虫(Autolytus),此类动物具有较强再生性,前半部分虫体可以脱离端细胞生长的方式再生出完整的后半部虫体。
沙蚕科、裂虫科(Syllidae)以及矶沙蚕科(Eunicidae)的不少种类具有一种特征性的生殖现象,称为生殖态,即在生殖时期通过整个个体的变态,或通过体后部体节的分化,使体后部体节发生显著变化,变为生殖节,是产生生殖细胞的体节;体前部体节基本保持原来的形态,不产生生殖细胞,称为无性节,此时身体明显分成两个不同的区域,也叫异沙蚕相,如沙蚕属物种在生殖态时,眼增大、特定体节的附肢增大、生殖节出现、肛门附近用于排出精子的器官发育。有这种现象的底栖种类在变态完成后,在受到光信号变化的影响时会同时(同一种)从海底游向海水表面,雌雄虫各自释放生殖细胞(雄性由雌虫产生的外激素刺激而释放精子),这种同步行为称为群游现象。
寡毛纲
寡毛类雌雄同体,异体受精,生殖系统常位于体前部,且生殖器官的不同部分各位于特定的体节内,雄性精巢两对或一对,雌性卵巢一对,受精囊一至数对,有几种水栖种类无受精囊。精子和卵在同一个体内非同时成熟。发育时间根据种和环境条件的不同可以从一周到几个月。
寡毛类性成熟时在一定体节上会形成环带(生殖带),所在体节数在陆栖种类中有所差异,在水栖种类所占体节数和位置各不相同。两个体交配时前端腹面相对,借生殖带分泌的粘液紧贴在一起,各自的熊生殖孔紧贴对方的受精囊孔,释放精液后分开,待卵成熟时,生殖带分泌大量黏稠物,于生殖带周围形成革质的蛋白质套管,管内有由生殖带分泌的大量白蛋白,卵排于其中,此后蚯蚓身体向后退,含有卵的蛋白质套管相对前移,当其移至受精囊孔时,受精囊内的精子排入套管内,在白蛋白的基质中与卵结合成受精卵,最后蚯蚓前端完全退出套管,管留在土中,两端封闭,形成卵茧,每个卵茧内受精卵数目根据种类不同而异,从1到约20个,然而仅有一个或少数达到孵化阶段,卵茧的形状和大小因种类而异,受精卵在卵茧内直接发育。此外,无性繁殖在水栖寡毛类中相当常见,如顠体虫属(Aeolosoma)能用无性的横分裂法繁殖。

交配中的陆正蚓(Lumbricus terrestris)
蛭纲
蛭纲动物全部行有性生殖,雌雄同体,经交配异体受精,具有生殖带,生殖季节常在春季,此时生殖带明显,交配受精后形成卵茧,受精卵在卵茧内直接发育。蛭类无受精囊,交配时两个体雌雄生殖孔彼此相对,借助阴茎将精子注入对方生殖孔,少数种类没有阴茎,而是将精荚注入对方生殖带的皮下,然后精子到卵巢囊内与卵结合。由于种类的不同,受精卵的数量从1~100个不等,这些受精卵通常被包裹在卵茧内,卵茧附着在岩石或植被上,受精卵约经一年成熟,成体寿命2~5年。
物种保护
编辑物种现状
截至2024年,根据《世界自然保护联盟(I.cn)濒危物种红色名录》,环节动物大部分种群数量变化趋势未知,在已知变化趋势的种类中,极少数种类呈上升趋势(1种,Amynthas noninvisus),大部分种类为平稳(30种,如丘疹远环蚓 Amynthas papulosus),其余种类呈下降趋势(9种,如帕卢斯巨型蚯蚓 Driloleirus americanus)。
致危因素
栖息地破坏
人类开发土地建成住宅、频繁的农业和水产养殖业是造成环节动物栖息地减少的主要因素,其次,矿业开采导致的地质变化也对环节动物的栖息地造成了破坏,但由于矿业开采的范围小于房地产业和农业、水产养殖业,故影响的物种数量少于后者。
如日本远环蚓(Amynthas japonicus)在被发现的两个世纪中由于日本的城市化和工业化导致了大量栖息地的丧失;Aporodrilus mortenseni 栖息地之一的帕默斯顿北部城市,人口从1900年的6000人增加到1916年的12000人,再到2016年的83500人,人口的大量增长伴随的是原生森林的清除和城市边缘的扩张,这些严重减少了Aporodrilus mortenseni 栖息环境。
种间影响
部分环节动物在栖息地的食物链中所占的营养级较低,容易受到入侵物种的影响导致种群数量减少,如天敌的入侵或同营养级入侵物种对食物等环境因子的挤占。
Anisochaeta animae 在特帕基的栖息地中一般会被野猪、老鼠、刺猬和负鼠等动物所捕食,外来的画眉对Anisochaeta animae的捕食加重了其生存压力;同时阿根廷蚁(Linepithema humile)的向北扩张对Anisochaeta animae 种群造成了更严重的威胁,二者的栖息环境和食物来源基本相同,除阿根廷蚁外,达尔文蚁(Doleromyrma darwiniana)种群也在逐渐向北扩张,其习性与Anisochaeta animae 同样相似,且Anisochaeta animae 仅在特帕基这一个栖息地被发现,所以二者的入侵对其来说是一个更加严重的威胁。
自然灾害
栖息地自然环境的剧烈变动以及自然灾害的影响会对栖息地狭窄或某地特有物种的种群数量造成极大的破坏:克马德克火山蠕虫(Acanthodrilus kermadecensis)所在的拉乌尔岛是一座活火山,在过去3600年中有16次喷发,其中4次发生在1814年、1870年的近代,在1949年发现到这种蠕虫后,该火山在1964年、2006年和2016年分别喷发,其中2006年3月的一场喷发导致半个岛屿都被火山灰覆盖,火山中喷出的气体、泥浆和巨石导致火山喷发中心的湖泊水位上升了6~8米,这种栖息地的剧烈变化会导致该蠕虫的种群数量在短时间内大幅降低,并伴随着无法恢复的风险。
保护级别
2023年,中国《有重要生态、科学、社会价值的陆生野生动物名录》中,巨蚓科(Megascolecidae)的参状远盲蚓(Amynthas aspergillus)、栉盲远盲蚓(Amynthas pectiniferus)、威廉腔蚓(Metaphire guillmi)、通俗腔蚓(Pheretima vulgaris)被列入其中。
2023年,环节动物门的医蛭科(Hirudinidae)的欧洲医蛭(Hirudo medicinalis)和侧纹医蛭(Hirudo verbana)被列入《濒危野生动植物种国际贸易公约(CITES)》附录Ⅱ。
截至2024年,环节动物门下共有359个物种被列入《世界自然保护联盟(IUCN)濒危物种红色名录》中,其中已灭绝(EX)的有2种,极危(CR)有8种,濒危(EN)有16种,易危(VU)有12种,近危(NT)有20种,无危(LC)有108种,数据缺乏(DD)有193种。
| 学名 | 状态 |
| Hypolimnus pedderensis | 灭绝(EX) |
| Tokea orthostichon | |
| Aporodrilus mortenseni | 极危(CR) |
| Arraia nelmae | |
| Eisenia anzac | |
| Maoridrilus felix | |
| Mesonerilla prospera | |
| Octochaetus levis | |
| Octochaetus microchaetus | |
| Perionychella ngakawau |
保护措施
环节动物因栖息地特殊性(土壤中或海底),尚无有效的保护区建立,但各地通过对入侵物种的查杀以及对原生林地、草地等的保护,一定程度上缓解了环节动物受到的威胁;相关的保护法也尚未健全,但对环节动物的保护意识依然在各地通过宣传教育的方式传递。
价值
编辑食用价值
多毛纲中的疣吻沙蚕(Tolorrhynchus heterochaeta)在中国广东被称作“禾虫”,被采食于稻田;沙蚕在中国其他地区也有多种烹饪方式;在福建沿海一带,沙蚕被称为“沙虫”,制成食物“土笋冻”。
寡毛纲中的蚯蚓在古代中国的广东省和福建省所在地区被作为食物,即便到现在,中国广东省、海南省和台湾省依然有将蚯蚓作为食物的现象;老挝、暹罗和日本等东南亚和东亚国家,食用蚯蚓的现象都是普遍存在的;在伊朗,蚯蚓也被烘烤并连同面包一同食用,当地人认为这样可以减少膀胱结石;毛利人会将蚯蚓制作成8种不同的菜肴(Kuharu、Noru、Wharu、Tarao、Pokotea、Tai、Kurekure、Whiti),其中Kurekure和Whiti因风味独特而闻名,并且被用来献给酋长或作为垂死之人最后的食物;澳大利亚原住民将蚯蚓作为额外的食物来源;在巴布亚新几内亚,萨卢姆河沿岸的游牧民族也会食用蚯蚓;委内瑞拉的一些部落会将蚯蚓作为美食,给分娩一个月后的妇女食用。
蛭纲动物在中国也有与生淮山一同煲制成粥的食用方法。
水产饵料
多毛纲的沙蚕和寡毛纲的蚯蚓作为钓鱼饵料被频繁使用;多毛纲的大多数种类为鱼类和很多海产动物的重要饵料,沙蚕的担轮幼虫为对虾幼体的优良食物;寡毛类的水栖种类可作为淡水鱼类的饵料,蚯蚓含有丰富的蛋白质,其含量占干重的50%~65%,含18~20种氨基酸,其中10种是禽畜必需的,作为动物性蛋白饲料,对家禽、家畜、鱼类及珍稀水产动物增产效果明显,如中国市面上出售的赤子爱胜蚓(Eisenia foetida)和美国红蚓杂交所得的“大平二号蚓”。
医用价值
蚯蚓入药在中国已有4000多年历史,《本草纲目》称其“治大腹黄疸”“治中风疾病”“疗伤寒”;中国现存可入药蚯蚓有参环毛蚓(Pheretima aspergillum)、威廉环毛蚓(Pheretima guillelmi)等,干制品即为中药的“地龙”,有清热息风、平喘、通络、利尿等功能,主治热病惊狂、小儿惊风、咳喘、头痛目赤、咽喉肿痛、小便不通、风湿关节疼痛、半身不遂等症;在公元1340年,印度和缅甸的医生就开始利用蚯蚓治疗一些疾病,如缅甸医生将蚯蚓在封闭容器内烧至灰烬后用以治疗一种伴有发烧症状的疾病,或与罗望子和槟榔等共同使用治疗一种导致产妇产后虚弱的疾病,印度则是在阿育吠陀医学体系中就存在着利用蚯蚓的营养成分治疗疾病的记录。
1991年,Mihara博士和日本的其他科学家成功地从红正蚓(Lumbricus rubellus)体内提取并表征了一组纤维蛋白溶解酶,即蚓激酶,对底物的特异性较强,故不会因蛋白质大量溶解导致组织大量出血,被用于溶解血栓和抑制血栓形成。
在埃及,使用水蛭进行治疗可以追溯到公元前1500年初。截至2024年,水蛭依然被用于治疗各种疾病,如皮肤病、关节炎和癌症。医用水蛭的唾液腺涉及100多种生物活性蛋白,唾液腺分泌物含有抑菌、镇痛和抗凝作用,通过分解能力消除微循环的障碍,恢复器官和组织的受损血管通透性、消除缺氧、降低血压并通过抗氧化产生解毒效果。在外科手术中通过医蛭吸血乐以便静脉血管通畅,减少坏死,提高手术成功率;中国于1987年首次用日本医蛭治疗断指再植手术后的淤血。
动物演化研究
多毛纲动物的幼虫阶段即担轮幼虫,不仅发生于环节动物,也存在于其他一些动物类群,对探讨动物演化关系有重要意义;环节动物在系统发生树中与软体动物门、星虫动物门等在同一分支上,在研究胚层的发展、体腔的变化等结构机制的进化上不可或缺。
环境管理
生活在土壤中的寡毛类动物因其吞吃土壤排出蚓粪的特性,可改变土壤的理化性质,增强土壤吸水、保水和透气性;蚓粪中含大量有机酸和腐殖质以及多种微量元素、氨基酸,可作为全价复合肥料;蚯蚓可用于处理造纸厂的污泥、酒厂的废弃物以及城市垃圾,还有聚集土壤中一些重金属的能力,英美等国在重金属矿附近的耕作区和农药污染的农田放养蚯蚓以减少和清除污染。
通过检测蚯蚓体内富集的金属含量可监测地区污染指数。
参考资料
编辑展开[1]Annelida.ITIS. [2024-03-24].
[2]刘凌云, 郑光美主编. 普通动物学[M]. 北京, 2009: 161-189. (9)
[3]Annelida.marinespecies. [2024-03-24].
[4]Annelida.IUCN. [2024-03-24].
[5]张士璀, 何建国, 孙世春主编. 海洋生物学[M]. 青岛: 中国海洋大学出版社 , 2017.12: 118-121. (5)
[6]环节动物门.中国大百科全书. [2024-03-24].
[7]普通动物学 第4版. 2009: 161-189. (29)
[8]王国秀, 闫云君, 周善义主编. 动物学[M]. 武汉: 华中科技大学出版社 , 2019.01: 105、109、110、115~117. (6)
[9]夏征农主编. 辞海 生物学分册[M]. 上海: 上海辞书出版社, 2019: 766.
[10]Zhang P, Misra S, Guo Z, Rehkämper M, Valsami-Jones E. Stable isotope labeling of metal/metal oxide nanomaterials for environmental and biological tracing[J]. Nat Protoc, 2019 : 2878-2899. [2024-03-24].
[11]Shakouri A, Wollina U. Time to Change Theory; Medical Leech from a Molecular Medicine Perspective Leech Salivary Proteins Playing a Potential Role in Medicine[J]. Adv Pharm Bull, 2021 : 261-266. [2024-03-24].
[12]Struck TH, Schult N, Kusen T, Hickman E, Bleidorn C, McHugh D, Halanych KM. Annelid phylogeny and the status of Sipuncula and Echiura[J]. BMC Evol Biol, 2007 : 57. [2024-03-24].
[13]The polychaete-to-clitellate transition: An EvoDevo perspective.ScienceDirect. [2024-04-07].
[14]Butterfield NJ. Exceptional fossil preservation and the cambrian explosion[J]. Integr Comp Biol, 2003 [2024-03-24].
[15]居峰, 董丽娜编. 紫金山动植物资源[M]. 南京, 2022: 249.
[16]大连近海无脊椎动物. 2017: 77、79. (2)
[17]Search Britannica.britannica. [2024-03-24].
[18]Annelida.GBIF. [2024-04-07].
[19]无脊椎动物学. 1961: 155.
[20]山东省水产学校主编. 海洋浮游生物学[M]. 北京: 中国农业出版社 , 1993.05: 142.
[21]Jumars PA, Dorgan KM, Lindsay SM. Diet of worms emended: an update of polychaete feeding guilds[J]. Ann Rev Mar Sci, 2015;7: 497-520. [2024-03-24].
[22]赵汝翼, 路顺奎编. 无脊椎动物学[M]. 北京: 高等教育出版社 , 1958.12: 187-188. (2)
[23]Kwak HJ, Kim JH, Kim JY, Jeon D, Lee DH, Yoo S, Kim J, Eyun SI, Park SC, Cho SJ. Behavioral variation according to feeding organ diversification in glossiphoniid leeches (Phylum: Annelida)[J]. Sci Rep, 2021[2024-03-24].
[24]Ribeiro RP, Egger B, Ponz-Segrelles G, Aguado MT. Cellular proliferation dynamics during regeneration in Syllis malaquini (Syllidae, Annelida)[J]. Front Zool, 2021 : 27. [2024-03-24].
[25]Amynthas japonicus.IUCN. [2024-03-24].
[26]Aporodrilus mortenseni.IUCN. [2024-03-24].
[27]Anisochaeta animae.IUCN. [2024-03-24].
[28]Acanthodrilus kermadecensis.IUCN. [2024-03-24].
[29]有重要生态、科学、社会价值的陆生野生动物名录.中国政府网. [2024-03-24].
[30]濒危野生动植物种国际贸易公约(CITES).国家林业和草原局 国家公园管理局. [2024-03-24].
[31]慈溪市政协教科卫体文化文史和学习委员会编, 邱雄飞著. 三北靠海文化田野调查[M]. 宁波: 宁波出版社 , 2021.08: 29-30.
[32]刘立身著. 闽菜史谈[M]. 福州: 海风出版社 , 2012.08: 174.
[33]Cooper EL, Balamurugan M, Huang CY, Tsao CR, Heredia J, TommaSEO-Ponzetta M, Paoletti MG. Earthworms dilong: ancient, inexpensive, noncontroversial models may help clarify approaches to integrated medicine emphasizing neuroimmune systems[J]. Evid Based Complement Alternat Med, 2012: 164152. [2024-03-24].
[34]卢俊主编, 陈东旭, 易腾编著. 常见病食疗手册[M]. 广州: 羊城晚报出版社 , 2003.09: 460.
[35]周希澄主编. 常用生物科技词典[M]. 郑州: 河南科学技术出版社 , 1989.02: 362.
[36]刘灿, 刘伟编著. 身痛逐瘀汤[M]. 北京: 中国医药科技出版社 , 2013.01: 164.
[37]林余霖编著. 本草纲目原色图谱800例 4[M]. 北京: 华龄出版社 , 2020.01: 716.
[38]Wang KY, Tull L, Cooper E, Wang N, Liu D. Recombinant protein production of earthworm lumbrokinase for potential antithrombotic application[J]. Evid Based Complement Alternat Med, 2013;2013: 783971. [2024-03-24].
百科词条作者:小小编,如若转载,请注明出处:https://glopedia.cn/82768/